
The Biological Effects of Ergothioneine
Explore the multifaceted biological effects of Ergothioneine, including its antioxidant, anti-aging, anti-inflammatory, and protective roles in skincare and health.
Antioxidant Effects
Ergothioneine (EGT) exhibits strong antioxidant capabilities even at very low concentrations (nanomolar) [50]. It can also neutralize various reactive oxygen and nitrogen species, as well as free radical generators, including hydroxyl [51], superoxide [51, 52], DPPH [53-55], ABTS [53, 55], and peroxynitrite radicals [51].
As an essential trace element, copper, along with other transition metals like zinc and iron, plays a crucial role in the active centers of various metalloenzymes involved in redox reactions [56, 57]. Copper is also linked to DNA bases, forming a critical component of chromatin and genetic material [58]. Abnormal copper levels can induce oxidative damage to DNA, proteins, and lipids, leading to apoptosis and pathological changes [59-62]. In the antioxidant network, most components undergo the Fenton reaction with copper, producing additional ROS and free radicals, which complicates achieving optimal antioxidant effects. Interestingly, EGT can rapidly chelate metal ions like iron and copper, forming stable complexes without undergoing the Fenton reaction [29]. Moreover, EGT competes with other metal complexes (such as histidine) for copper ions, significantly inhibiting the formation of protein carbonyls and thus protecting DNA and proteins from copper-induced oxidative damage [63-65].
The exact role of EGT in the antioxidant network remains unclear; however, studies suggest that EGT enhances the stability of other antioxidants and even acts synergistically within the body. Gregory et al. [66] demonstrated EGT’s superior antioxidant properties using three antioxidants (EGT, ferulic acid, and GSH). When combined with 100 μM ferulic acid, 50 μM EGT showed enhanced antioxidant capacity, preventing oxidative damage and photoaging in skin cells. Additionally, low concentrations of EGT significantly increased the clearance rate of the DPPH radical by GSH and niacinamide. Numerous studies have shown that EGT significantly raises GSH levels, accelerates the GSH/GSSG cycle, and promotes the reduction in GSH production [67]. EGT also supports the multi-step non-enzymatic cycles for singlet oxygen removal and works in conjunction with glutathione, indicating that higher glutathione levels allow EGT to regenerate after being consumed by singlet oxygen. Notably, EGT reacts with singlet oxygen much faster than vitamin C and L-histidine [20]. EGT’s stability and reusability often make it a stabilizer to prevent discoloration and spoilage, exhibiting strong antioxidant effects in vitro, delaying the aging and death of tissue cells [68, 69].
Additionally, EGT helps maintain and regulate NADPH levels, an essential component of the cellular antioxidant system and a source of electrons needed for fatty acid, steroid, and DNA synthesis [70, 71]. However, when NADPH is oxidized to NADP by NADPH cytochrome c reductase, it produces H2O2. Also, under the action of the NADPH oxidase (NOXs) family of enzymes, substantial amounts of ROS are generated, a significant source of mitochondrial ROS [72]. Ideally, EGT can chelate metal ions that promote H2O2 reactions and directly inhibit NOXs activity, thereby reducing the pathways that generate ROS [8].
Anti-Aging Effects
The skin, being the largest organ and outermost barrier of the human body, is the most visually evident indicator of aging influenced by age and external factors [73]. The process of skin aging is complex, generally attributed to exogenous and endogenous aging [74]. Exogenous aging involves damage or premature aging of cells due to environmental factors such as ultraviolet radiation and pollution [75]. In contrast, endogenous aging reflects the decline in cellular immune defense mechanisms [76] and self-renewal capabilities with age [77]. Depletion of stem cells and diminished metabolic capacity lead to the skin cells’ inability to renew and manage harmful substances, ultimately causing skin aging. During this process, mitochondrial nicotinamide adenine dinucleotide (NADH) and NADPH play key roles in the cellular antioxidant system, energy metabolism, and electron transport. Furthermore, mitochondria are responsible for approximately 90% of cellular energy production through oxidative phosphorylation, generating substantial amounts of ROS [79, 80]. When ROS accumulate abnormally, the body can regulate ROS levels through its antioxidant defense systems. However, some ROS may bypass these processes, potentially damaging mtDNA, proteins, and lipids. Through these mechanisms, cellular aging can be delayed, and repair capabilities enhanced [79].
NADH is primarily stored in mitochondrial NADH pools, involved in cellular energy metabolism [81]. OCTN-1 expression is also high in mitochondria, suggesting EGT’s significant role in mitochondria as shown in Figure 2. EGT can regulate NOXs [82, 83], NADH oxidases [31, 80, 84], and GSH-Px [84]. Thus, it is hypothesized that OCTN-1 specifically transports EGT to mitochondria, clearing intracellular ROS and alleviating the aging process. Additionally, some studies have confirmed that EGT affects NADH oxidases [31], promoting the conversion of NADH to NAD+ stored in mitochondrial pools, ultimately lowering NAD+/NADH levels [85]. This suggests that EGT reduces the potential generation of ROS and activates SIRTs [71, 85], PARP-1 [33, 71], which are significant.
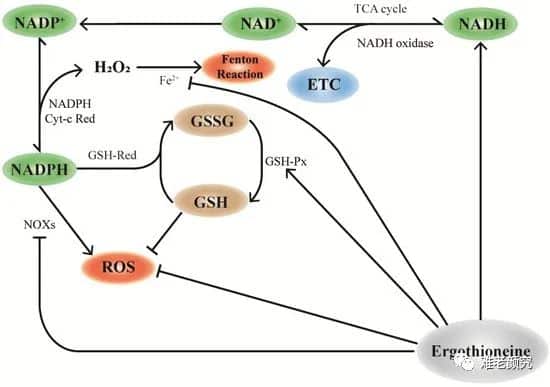
Figure 1. Mechanism of EGT and NAD+ recycling pathways. In mitochondria, the consumption of NAD+ participates in the TCA cycle of cellular energy metabolism. The NADP+ produced by NAD is affected by NAD kinase and further generates NADPH. NADPH serves as a substrate for glutathione reductase (GR), which reduces GSSG to glutathione (GSH), the antioxidant enzyme glutathione peroxidase (GPx) and glutathione transfer Enzyme (GST) is required for activity and is one of the important ways to improve the antioxidant capacity of cells. EGT promotes this effect by modulating the action of NAD+ pools in mitochondria. In addition, the antioxidant properties of EGT can directly remove excess ROS, increase the activity of GSH-px, and synergistically participate in the antioxidant effect of cells.
As shown in Figure 1, hypotheses and inferences can be made about the maintenance and regulation of GSH by EGT. According to research, EGT can inhibit the activity of NADPH oxidase, thereby reducing ROS produced by NADPH oxidation, increasing the level of NADPH, and maintaining the reduced state of GSH [14]. In addition, NADPH can maintain the stable state of GSH under the action of GSH reductase, generate GSSG through GSH red oxidase, and decompose H 2 O 2 with the participation of EGT. The increase in GSH results in a corresponding increase in the GSH/GSSG ratio [ 31 ]. Pan et al. [83] confirmed that EGT can maintain the GSH/GSSG ratio and extend lifespan in Drosophila.
Anti-Inflammatory Effects and Immune Modulation
Oxidative stress or environmental factors often trigger immune system stress, leading to inflammatory responses and pathological changes [86]. Among these factors, exogenous elements such as ultraviolet radiation stimulation [87], harmful gases [88], and trauma [89] are common, triggering inflammation pathways and activating pro-inflammatory factors like IL-1β, IL-6, IL-8, and TNF-α [90]. Besides directly causing cell and tissue damage, these factors also activate other cells in the skin, such as Langerhans cells and lymphocytes [91], releasing inflammatory mediators [92], causing local reactions, and inducing dermatitis and other skin diseases, including skin cancer [93]. Therefore, researching how to mitigate the inflammatory process or block inflammation pathways to maintain skin barrier and health is crucial.
As a component of the human antioxidant system, EGT has been studied as a potential anti-inflammatory agent, suggesting it acts as a backup for accumulating antioxidants in the body and specifically functions at damaged sites [27, 94, 95]. Understanding the role of EGT signal transduction in cellular oxidative stress is essential. As shown in Figure 3, EGT likely activates the body’s antioxidant repair systems by regulating cellular factors like TNF-α, NF-κB, IL-6 through the activation of intracellular MAPK gene cascades [27, 96]. Additionally, it modulates the activity of MMP-1. NOXs [8] and other inflammation-related enzymes [50]. As shown in Figure 1, when the body is stimulated by factors like H2O2, UV, PA, OCTN-1 is rapidly expressed or transmitted, significantly increasing the EGT content at damaged sites [96].
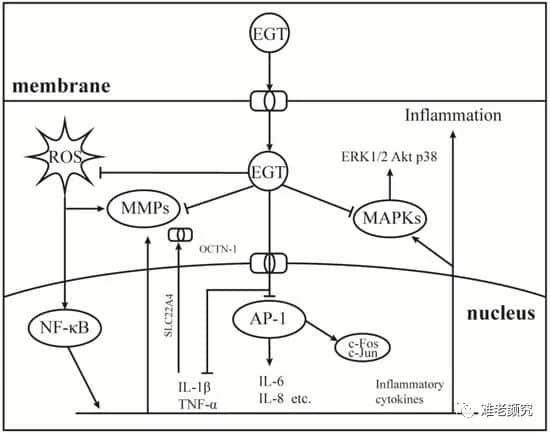
Figure 2. Anti-inflammatory mechanism of EGT on AP-1 and MAPKs signaling pathways. Induced by environmental stress, ultraviolet light, or oxidative damage, ROS can easily attack the cell nucleus and produce a large amount of inflammatory cytokines, thereby activating the NF-κB cascade signaling pathway. Therefore, the cell nucleus begins to express a large amount of OCTN-1 to promote the transport of more EGT into the cell to play its role. While scavenging ROS, EGT inhibits the production of activated AP-1 and MAPK cascade signaling pathways, thereby alleviating inflammation.
Furthermore, EGT stimulates the inhibition of TNF-α and activates the NF-κB, inducing the protective effects of the ARE/Nrf2 and Hsp70 pathways, activating the antioxidant system, rapidly clearing and suppressing the production of harmful free radicals and inflammation factors [50]. Additionally, as EGT accumulates, oxidative damage and inflammatory markers (such as orosomucoid, 8iso-PGF2G, and C-reactive protein) at stress sites significantly decrease [8]. Interestingly, the expression of OCTN-1 may lag behind the onset of inflammation, and OCTN-1’s gene expression region is located on chromosome 5q31, associated with multiple immune and inflammatory system genes [95]. This may imply that OCTN-1 and EGT are closely related to the immune system, serving as the body’s last barrier against damage.
High expression of OCTN-1 in immune cells also indicates EGT’s role in immune regulation. It has been demonstrated that EGT may act as a Toll-like receptor agonist involved in immune system regulation [97]. EGT specifically acts on macrophages under the participation of OCTN-1 transport proteins, inducing the transformation of CD4+ T cells into Th17. Additionally, EGT pretreatment enhances the transcription of M1-related cytokine genes induced by TLR ligands, relatively reducing IL-10 expression. It is well-known that IL-10 is often associated with inflammatory lesions of acne and suppresses the activation and proliferation of T cells by inhibiting antigen-presenting cells such as Langerhans cells [98]. Additionally, EGT is involved in suppressing JNK-mediated IL-6 and TNF-α-mediated IL-8 production [96, 99], playing a key role in immune and inflammatory responses. However, the role of EGT in immune cells may differ from its function in other cell types, as evidenced by EGT acting as a TLR agonist, enhancing the expression of IL-1β, IL-6, and IL-12p40. IL-6 and IL-23 regulate IL-12p40 to promote Th17 differentiation in naive CD4+ T cells [97]. Although the antioxidant effects of IL-6 and IL-12p40 production are controversial, the degree of Th17 polarization induces the promotion of inflammation-based autoimmunity.
Protection of Skin Barrier
The skin consists of the epidermis, dermis, and subcutaneous tissue, forming a complete barrier. These layers are differentiated and arranged by various cells, constituting the first interface between the body and the environment, as shown in Figure 4. Additionally, the skin forms a physical, chemical, and stable biological barrier that can accommodate different microorganisms and maintain ecological balance. Damage to this skin barrier function may lead to skin diseases or infections, such as atopic dermatitis (AD) and acne. Rizzo et al. [100] demonstrated that the p53 family member p63 could be a key driver of AD. Furthermore, p53 affects cell proliferation and differentiation by regulating RUNX1, which is usually detected in the nuclei of epidermal cells but is not significantly expressed in AD patients [101]. This may suggest a connection with the reconstruction of skin structure and the repair of the skin barrier.
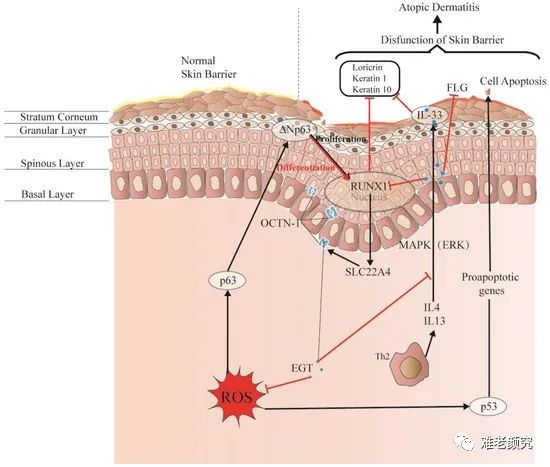
Figure 3. EGT may regulate RUNX1 through the protective skin barrier. In addition, EGT can also downregulate the inflammatory factor IL-33 induced by MAPK kinase, reduce the production of inflammatory markers (loricrin, keratin 1 and keratin 10), and indirectly regulate apoptosis caused by p53/63 by inhibiting the excessive increase in ROS. Interestingly, the expression of EGT and RUNX1 proteins induces strong binding energy associated with the proliferation and differentiation of keratinocytes or skin stem cells, and ultimately promotes the renewal of skin barrier function.
Interestingly, the expression of OCTN-1 is strongly correlated with RUNX1 gene expression (score=0.999), as reflected in the STITCH database, as shown in Figure 5. Additionally, a study indicated that EGT has a positive effect on immune system diseases caused by abnormal RUNX1 expression, such as rheumatoid arthritis (RA) [102-104] and non-alcoholic fatty liver disease [12]. Additionally, the expression of RUNX1 is similar to that of p63 and p53, directly regulating cell proliferation and differentiation processes, which is also an important regulator of normal hematopoiesis in the body, possibly consistent with the high concentration results in the blood. EGT [101]. Additionally, it is a key regulator of normal hematopoiesis [105], which may be consistent with the abundant accumulation of EGT in the blood [15]. Furthermore, RUNX1’s regulatory effect on SLC22A4 and ROS’s activation of p53 may suggest that EGT, through high expression of OCTN-1 transport, clears excess ROS, or the anti-inflammatory action of IL-33 alleviates AD after AD. Skin barrier function is impaired. This may lead to the downregulation of IL-33-mediated inflammatory markers (involucrin, keratin 1, and keratin 10) [106]. Overall, these findings suggest that EGT may play an important role in reshaping skin barrier function and treating AD.
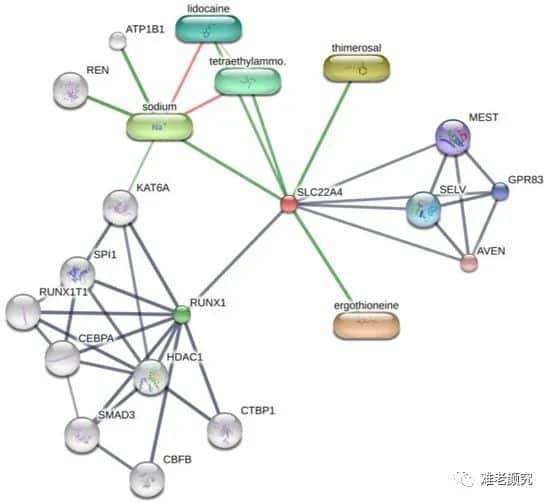
Figure 4. Prediction of EGT binding to related proteins by STITCH database cluster analysis.
Protection from Ultraviolet Radiation
Ultraviolet radiation is one of the most significant factors causing skin damage and aging [107]. It is well known that under ultraviolet stress, photoreactions in cells or tissues form many harmful products, causing skin damage, such as the formation of ROS, including superoxide anion, singlet oxygen, hydrogen peroxide, and hydroxyl radicals [108]. It can induce a series of MAPK pathways and produce inflammation [27, 109]. Therefore, as an important component of the human antioxidant system, EGT will undoubtedly become a research hotspot for potential skincare products. Additionally, EGT has an absorption spectrum within the ultraviolet range, with a molar extinction coefficient of 1.4 × 104 M-1·s-1, and a maximum wavelength of 257 nm [10], similar to common sunscreen octinoxate (320 nm) [110]. The absorption band at 258 nm may result from the electronic transition of the EGT 2-thioimidazole part in the thione form, which is disrupted by the loss of nitrogen protons (pKa 10.8) [111]. This suggests that the thione state of EGT can act as a physiological ultraviolet filter, absorbing or shielding ultraviolet radiation and producing biological active effects, which may be one of the reasons it prevents damage caused by ultraviolet radiation. However, ultraviolet-induced decreases in skin collagen synthesis and matrix metalloproteinase activity are characteristics of photoaging, causing roughness, wrinkles, yellowing, capillary dilation, pigment loss, and even skin cancer, among other symptoms [25]. In vitro, EGT has been proven to be an effective inhibitor of AGE formation and AGE-collagen cross-linking [24], and by reducing MMP-1 degradation and enhancing type I collagen production, it protects these cells [79]. This means that EGT, as a natural antioxidant and effective anti-photoaging agent in the skin, has many advantages and has the potential to become an excellent ultraviolet protective agent for developing skincare products or healthy sunscreens [79, 112].
The mechanism of EGT’s ultraviolet protection is similar to that of antioxidants and inflammation reduction. This means that ultraviolet radiation induces and exacerbates oxidative or inflammatory damage. However, the difference is that prolonged ultraviolet radiation exposure may cause additional DNA damage. Studies have shown that the characteristic of photoaged skin is an increase in mitochondrial genome mutations [113]. The most common deletion is a large-scale (4977 bp) deletion, also known as the “common deletion” (CD) [114, 115]. In human skin, as ultraviolet radiation exposure increases, the frequency of CD occurrence also increases, 10 times more in photoaged skin compared to volunteers using sunscreen [114]. Based on the mechanism of action of EGT and the distribution of OCTN-1, it can be inferred that EGT acts on mitochondria to inhibit DNA damage. Additionally, a study showed that when 20 µM EGT was co-incubated in fibroblasts exposed to UVA for three weeks, no CD was observed, indicating that EGT can prevent this specific damage in mtDNA [79]. Another study showed that 500 nM EGT pretreatment effectively prevented UVA-induced TUNEL-positive nuclei and mitochondrial membrane potential loss [50]. Therefore, EGT may be involved in protecting mitochondrial DNA during the electron transport cycle from adverse effects.
Inhibition of Melanin
Melanin in the skin serves as a natural barrier against ultraviolet protection, playing a crucial role in maintaining skin health [116]. However, excessive or insufficient melanin can lead to abnormal skin color and skin lesions, such as age spots and melasma [118, 119]. Therefore, skincare products with whitening and spot-removing functions occupy an important position in cosmetics. Recently, many studies have focused on the activity of tyrosinase and the changes in the amount of melanin produced by cells or tissues.
The mechanism of EGT’s inhibition of melanin production or removal may be due to its unique structure, namely the sulfur-substituted imidazole ring [120, 121]. Sulfhydryl compounds are considered important melanin production inhibitors because they react with dopaquinone to form colorless adducts [122]. Additionally, the uniqueness of EGT lies in the presence of its sulfur atom in the thione form, rather than in the thiol form [123]. This means that the thione structure may distinguish EGT’s inhibitory mechanism from other tyrosinase inhibitors (such as thiols). Additionally, Liao et al. [120] demonstrated that EGT can bind to the free enzyme’s substrate site and the enzyme-substrate complex’s different substrate sites. Furthermore, observations on inhibitor concentration showed that EGT did not reduce the amount of enzyme but caused inhibition of enzyme activity, indicating that EGT is a reversible tyrosinase inhibitor [120]. Additionally, other studies have also confirmed EGT’s inhibitory effect on tyrosinase; this inhibitory effect is superior to common inhibitors such as kojic acid and arbutin [55, 124].
Applications of Ergothioneine in Cosmetics
In recent years, natural ingredients have received widespread attention in cosmetics. Consumers’ demand for cosmetics containing natural or organic ingredients is continuously growing, as these components are considered healthier, safer, and more ecological. Due to its excellent efficacy, stability, and water solubility, EGT is adaptable to most formulations. Additionally, EGT produced through biosynthetic technology has higher purity and sustainability than natural extracts. Table A2 summarizes some skincare products containing EGT (marketed in 2023). EGT has been widely applied in nutrition, anti-aging, and sunscreen cosmetics, and is not limited by specific formulation systems. Additionally, oral nutrition cosmetics or supplements containing EGT have been marketed, often interacting with other vitamin components. However, the application of high-purity EGT or fermentation products in cosmetics is relatively expensive, suggesting that more cost-effective synthetic purification technologies also have research value.
About ETprotein:
ETprotein, a reputable protein and L-(+)-Ergothioneine (EGT) Chinese factory manufacturer and supplier, is renowned for producing, stocking, exporting, and delivering the highest quality organic bulk vegan proteins and L-(+)-Ergothioneine. They include Organic rice protein, clear rice protein, pea protein, clear pea protein, watermelon seed protein, pumpkin seed protein, sunflower seed protein, mung bean protein, peanut protein, and L-(+)-Ergothioneine EGT Pharmaceutical grade, L-(+)-Ergothioneine EGT food grade, L-(+)-Ergothioneine EGT cosmetic grade, L-(+)-Ergothioneine EGT reference grade and L-(+)-Ergothioneine EGT standard. Their offerings, characterized by a neutral taste, non-GMO, allergen-free attributes, with L-(+)-Ergothioneine purity over 98%, 99%, cater to a diverse range of industries. They serve nutraceutical, pharmaceutical, cosmeceutical, veterinary, as well as food and beverage finished product distributors, traders, and manufacturers across Europe, USA, Canada, Australia, Thailand, Japan, Korea, Brazil, and Chile, among others.
ETprotein specialization includes exporting and delivering tailor-made protein powder and finished nutritional supplements. Their extensive product range covers sectors like Food and Beverage, Sports Nutrition, Weight Management, Dietary Supplements, Health and Wellness Products, and Infant Formula, ensuring comprehensive solutions to meet all your protein needs.
As a trusted company by leading global food and beverage brands and Fortune 500 companies, ETprotein reinforces China’s reputation in the global arena. For more information or to sample their products, please contact them and email sales(at)ETprotein.com today.










